Stephen’s research focuses on understanding the causes and consequences of natural selection, specifically in sexually reproducing populations. In this seminar, he will argue the following points: sexual dimorphism, or within species differences between the sexes, are a pervasive form of biodiversity, often with ecological importance. He will then present a series of experiments from salamanders and flies that test for a role of direct ecological causes of sexual dimorphism – that is, ecological character displacement between the sexes. He will then go on to explore the theoretical and realized consequences of sexual dimorphism for the assembly of ecological communities.
This post covers chapters 21 & 22 from Futuyma and Kirkpatrick’s book on Evolution (2018). The author of this post is Sam Shry.
Evolution and Homo sapiens; the birds and bees of where we come from
We now turn our attention to the evolution of our own species and how we came to be this unique, evolutionary phenomenon. Of course, we have already learned that we arose from a last universal common ancestor (LUCA), but closer in evolutionary time we arose from the group called great apes, more specifically our closest relatives are the chimpanzee (Pan troglodytes) and the bonobo (Pan paniscus). The lineage hominin (us) diverged from the chimpanzee lineage around 7 Mya, with Homo sapiens becoming the only species from this lineage not to go extinct. With genetic similarities in our protein-coding genes of more than 98% between chimpanzee and humans, it’s no wonder that there are such close similarities between these two species in terms of morphological, social, and cognitive features. We, however, differ in many aspects too, for example, we have evolved the ability to live upright on two feet (bipedal), adapting to a changing climate during this time via movement and migration. We also evolved reduced body hair to probably aid in making sweating and evaporative cooling more efficient. We also have many differences in our hand morphology, teeth, and our larger brain. What is interesting to me is that our potential growth rate is so much higher than for other ape species, as human females can become pregnant even when they have dependent children, in contrast to chimpanzees that cannot.
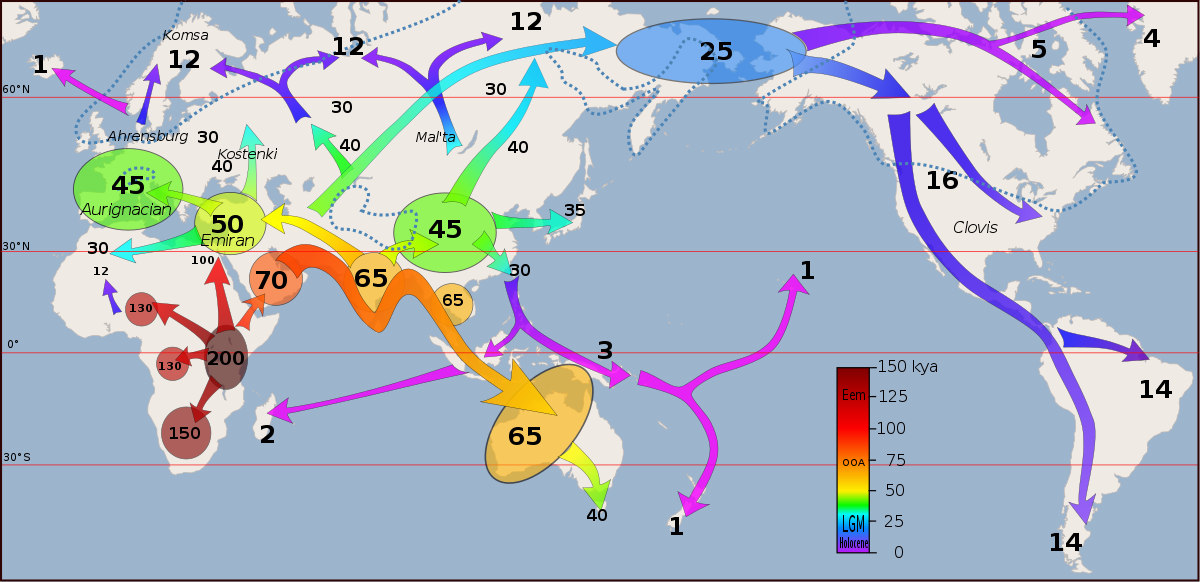
The divergence of hominins occurred in Africa, as Darwin predicted long before fossil evidence. Hominins split into several species, with all (except us) eventually dying out, giving an interesting insight into the evolution of our species. One of these (Ardipithecus ramidus) had apelike features like a chimpanzee’s brain size and climbing adaptations, but also had small canine teeth and a pelvis adapted for walking upright. The genus Homo dates back about 3 Mya, where the Homo habilis and Homo erectus are thought to be the ancestors of our own species. Homo habilis resembled modern humans closer than earlier species, with a flatter face, shorter tooth row, and greater cranial capacity than before. Homo erectus had a resemblance that was even closer to modern humans and was the first hominin to leave Africa around 2 Mya, spreading into the Middle East and eventually Asia. Our ancestors continued to slowly disperse across the world, with Homo heidelbergensis, a common ancestor to the well-known Neanderthals, dispersing throughout Europe and Asia around 600 Kya (figure 1). I like the analogy in the book that describes the phylogeny of hominins as a densely tangled bush, with humans only being a leaf on this bush.
Figure 1. Migrations and colonization of Earth by humans. (Wikimedia Commons, User:Dbachmann)
We can map our ancestral dispersal across earth over time using our gene trees of mitochondrial DNA (mtDNA) sequences to complement and provide additional insight to fossil evidence and estimate the age of the most recent common ancestor of mtDNA in living humans. We also find that these different groups of hominins hybridized, for example, Neanderthals and Denisovans, contributing advantageous alleles to the human gene pool. In today’s human populations, we also find some genetic differentiation, but these differences are very low compared to other species, due probably to the very short amount of time we have evolved since colonization. What is surprising is how adapted some human populations are to their environments due to high selective pressure, for example, lighter skin color in Europeans and East Asians adapted to limited sunlight in northern latitudes.
One important characteristic of humans is our brain size. Our cognitive abilities far surpass other species, giving us the ability to reason, think, and understand. One hypothesis for this is ecological, in that we evolved cognition for dealing with complex environments. Another hypothesis is called the social brain hypothesis, where our complex social groups selected for large brains that would ultimately increase survival and the chance for reproduction (increasing fitness). Social cooperation and accumulated knowledge via the use of language are key elements to the evolution of increased cognitive abilities in our species. The evolved modification of our vocal tract enables vocal versatility, enabling a wider range of sounds, but at the cost of possibly choking to death on food, so the advantages of spoken language must be strong in our evolution.
These somewhat strange evolutionary tradeoffs are further seen when looking at the costs of a larger brain, causing higher reproductive costs and requiring more energy to grow all the way up to 18 years, making human life history paradoxical. We do however have longer life spans than other primates, we evolved higher metabolic rates, higher reproductive rates, and a larger energy budget. We did this by switching diets to adding meats and tubers, and learning to hunt and gather. Agriculture was developed and animal domestication began to feed our growing population. Agriculture was double-sided, providing food stability, but changing human lifestyle, culture, and environment around us. Adapting to a new diet selected for specific genes, for example, the lactase persistence enzyme that allows for the digestion of lactose in milk. What is also interesting is how our ancestor dispersal from Africa left traces in our DNA sequence variation, where individuals that disperse furthest from Africa have the highest deleterious mutations, given they were small populations (founder effect), accumulating rare variants, less effective natural selection, and intense drift that fixed these mutations.
Natural selection and evolution are still acting on our species, it’s just that the selection pressures have changed. We find modern hygiene and medicine alleviate the fitness differences, but still, we see selection occurring in some traits, such as cholesterol levels and height, due to our diet, lifestyle, and environmental factors. We also still see the effects of changing to an agricultural diet in our obesity crisis, with calorie-dense foods combined with our sedentary lifestyles. We also have the evolution of culture acting on our society, simultaneously driving cultural elements, innovation, and social interactions. The cumulation of cultural influences, human behavior, and biological tendencies guide, but do not limit our diversity as we continue to move forward in time.
Evolution and Society
The theory of evolution by natural selection has had a profound impact on our understanding of ourselves and the world around us, “unifying the realm of life, meaning and purpose with the realm of space and time, cause and effect, mechanism and physical law.” – Daniel Dennett
Although there is a long line of evidence behind this theory’s confirmation, there are still those that do not accept this theory as reality and cannot grip general reasoning. An extreme, sub-population have “beliefs” in creationism, that God directly created the human species. Certain, literal interpretations of the Bible and other religious documents have conflicting ideas of Earth’s creation and its life, for example in the book of Genesis and the literal interpretation of the creation of Earth and life in six days. Others believe evolution is the mechanism by which God enabled/s creation to proceed and now lets the Earth run on its own without further intervention. People can believe whatever they want, but issues arise when religion and beliefs are confused with science and education. For example, in the US, some schools are required to give equal teaching time to religious doctrine like intelligent design as they do to scientific theory such as evolution. The problem with this is that people do not understand how science works and how it is not comparable to religion. The process of science is to forever test our understanding of the world through the scientific method. Over time we have come up with our best understanding of the world and these denoted theories are “proven” beyond a reasonable doubt via scientific testing and evidence-based science. As this book has explained over the last 21 chapters, the theory of evolution is a highly complex system that we are continuing to understand better every day. Evidence for evolution has been already described throughout the book but we can just recap a few examples. The fossil record and the transitional fossils between species have given us physical evidence of evolution that often matches up, time-wise, with phylogenetic predicted sequences. The phylogenetic relationship between species via DNA analysis has opened a vast new world of understanding every species connection and diversity. Though this evidence does not waver for some true believers, the failed argument of intelligent design gives a baseless ground for these beliefs. The whole argument of a godly design is that these complex species are created perfectly. So, the vast number of failed, inferior designed traits or species in this world would indicate an unkind, incompetent designer. Why would an ultimate designer create sickle-cell disease, saving some and killing everyone else that is homozygous for a gene? Intelligent design is unable to explain the selfish behaviors that natural selection can explain, like cannibalism, siblicide, and infanticide. We have also been able to use the information gleaned from the mechanisms governing the theory of evolution to understand the world around us and how it is changing. For example, understanding domesticated plants and animals, resistance to pesticides and antibiotics, and rapid adaptations to climate changes. In short, the evidence behind evolutionary theory should never be compared to beliefs and even if they were to be, there is no evidence for these beliefs.
Understanding evolution and its mechanisms has shaped our understanding of the world today. It has formed the world we live in, ourselves, and every other species. The concept of natural selection can be applied to all aspects of our lives, such as culture, language, political science, and the overall human experience. The practical application of evolutionary mechanisms has helped us in advancing food production, managing natural resources, improving conservation, and human health. By having this understanding of how organisms evolve, we can make future predictions for our species, other species, and even understand how to develop new technologies and organisms to further benefit us or save our species. This work is vital to combating today’s challenges such as climate change, biodiversity loss, genetic diseases, cancer, infectious diseases, and general public health. One example is the current extinction crises, the global loss of species diversity. Due to rapid climate change, species are unable to genetically adapt to new climate conditions, causing species extinction. Evolution is a unifying theme throughout the biological and social sciences and is the backbone to understanding the world around us. It is, however, only knowledge, with no external meaning, morals, or ethics. It is important knowledge for our society, one species of the millions inhabiting this earth. It is always fun to think about how little evolutionary time our species has inhabited this earth and how/if we will endure or just become another species on the extinction list.
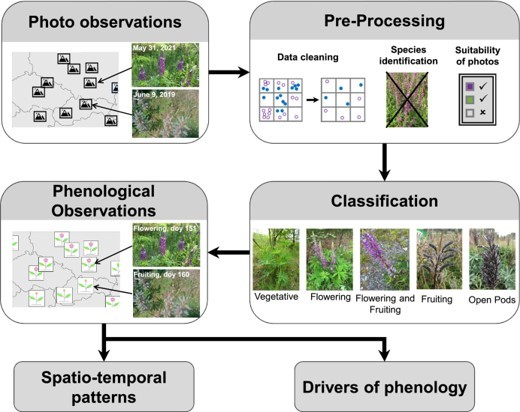
Lutz Eckstein is co-author of a paper presenting a workflow for iPhenology (Fig. 1), i.e., the use of publicly available photo observations to track phenological events at large scales. The paper, which has recently been published open access in the journal Methods in Ecology and Evolution (https://doi.org/10.1111/2041-210X.14114), is led by Yves Klinger (Justus-von-Liebig University Gießen, Germany), who will be visiting Biology at KaU in June 2023. Overall, publicly available citizen science photo observations are suitable to track key phenological events and can thus significantly advance the knowledge on the timing and drivers of plant phenology. In future, integrating the workflow with automated image processing and analysis may enable real-time tracking of plant phenology. To learn more about what sparked the idea for this paper and what may be advantages of working with citizen science photos, see the Methods in Ecology and Evolution blog (https://methodsblog.com/2023/05/22/using-citizen-science-photos-to-perform-phenological-studies/).
Figure 1. Proposed workflow for iPhenology. First, observations are pre-processed by removing problematic observation and, if necessary, reducing spatial aggregation. Second, photos are checked for correct identification and suitability before being classified. Unsuitable or misidentified photos are removed. For the resulting phenological observations, there are many potential uses.
This post covers chapters 8 & 9 from Futuyma and Kirkpatrick’s book on Evolution (2018). The author of this post is Jacqueline Hoppenreijs.
Evolution in space
Variation in phenotypic traits can reflect variation in environmental factors on a global scale, such as temperature, or on a local scale, such as nutrient or toxicant concentrations. When this phenotypic variation looks like a smooth transition or gradient, we call it a cline. Such ecological variation is a consequence of the selection pressure’s spatial variation and can be seen between and within species, leading to specific patterns of phenotypic traits. An example of such a pattern between species is the occurrence of hairs and cuticles in drought-adjusted plants, that help them retain small amounts of water that they have managed to get a hold of (De Micco & Aronne, 2012). A within-species example is the variation in root depth and structure in individuals of one species that are grown in drier circumstances than others (Lenssen et al., 2004).
Trait patterns are further shaped by exchange between populations of the same species, a process called gene flow that happens through dispersal. Dispersal is different from migration in that it means that an organism ventures out to explore new grounds (or waters) and that it leads to exchange or introduction of genes. Dispersal can be passive (e.g. a seed falls into the water and ends up on a riverbank elsewhere), or active (e.g. an animal starts looking for a more suitable living environment if it runs out of food). There are also forms that could be seen as intermediate, such as plants (Figure 1) that disguise themselves to convince someone else to fix their dispersal for them!
Figure 1. A dung beetle dispersing poop-disguised seeds from the species Ceratocaryum argenteum.
Dispersal can be a bit of a pain, as it requires energy (even if it’s only the effort of growing your seeds in the form of poop) and can be dangerous. It can have big advantages though: by dispersing, you get the opportunity to find a more suitable spot to live and you simultaneously decrease the day-to-day competition with your family members in the place where you all are from. You’re also more likely to reproduce with an individual from a different population, thus decreasing the risk of inbreeding.
So, it’s clear that dispersal can be very profitable, and leads to exchange of genes between populations or establishment of new populations. Gene flow can be measured in multiple ways, that are applicable in discrete (think: migration rate between islands) or continuous (think: migration variance throughout a vast forest) environments, or both. This can be your typical mark-recapture set-up, but more and more often genetics-based methods are used. There are many ways to go about this, but one straightforward approach is to assess genetic difference between populations using their allele frequencies, a variable that then can be combined with the populations’ physical distance to give an idea of gene flow history. Things become less straightforward when there’s local selection involved, that favours one allele over the other and thus (partially) undoes the effect of gene flow. Which of the two processes predominates depends on both local selection and migration rate/variance. If local selection were the dominant process, different populations would have different alleles that fit their environment, and if gene flow were the dominant process, populations would have the exact same allele, also called gene swamping. The two processes often keep each other in somewhat of a balance, and understanding them can help trace past events and processes, and predict future developments (e.g. Love et al., 2023).
Life as an evolutionary biologist is complicated enough with just these two processes going on, but there’s another one that comes into play: genetic drift. This process, that we heard about in blog 3 , is another source of genetic variation and it can be pretty complicated to find out whether it’s this or local adaptation (or both) that cause genetic and phenotypic differences between populations. Mapping of entire genomes makes it possible to look for differences between them, and can offer a solution to this problem. A comparison between the genetic variance in neutral and “under selection” regions in two or more different populations, gives you information on both. As the neutral regions will have a certain amount of variance that’s not a consequence of the environment favouring one allele over the other, this variance is most likely the consequence of genetic drift. The variance in the “under selection” region minus the variance in the neutral regions is than likely to be the result of local adaptation. It’s as simple as that!
Understanding local adaptations in the context of dispersal to new areas can open a whole new can of worms/research questions. One might wonder how other species in the region have adapted to their environment in the past and are now dealing with a new species entering the stage. Ecological research often has a competition-for-resources angle (Simha et al., 2022), but could for example also focus on niche construction (Laland et al., 2016). It’s thus virtually impossible that the entrance of a new species in an area will go unnoticed by the already present species. Researching this, however, requires somewhat of an understanding of how species are defined and come to be, which is our next topic.
Species and speciation
As we know now, species can adapt to new places that they disperse to, and to new circumstances that they’re confronted with. This can ultimately lead to the formation of one or more new species, also called speciation. Before understanding that process, it can be helpful to look at its results: how do we define species as different from each other? The answer is that this is almost always is arbitrary, and the approach can be simple: “they look different”. It gets a bit more complicated with “they can successfully reproduce”, also called the Biological Species Concept (BSC), or quite complicated with the approach “they are the smallest set of organisms that has the same ancestor” which is one of the versions of the Phylogenetic Species Concept (PSC). Why do I write “one of the versions”, you ask? That’s because there’s hardly ever a clear line to draw in biology: think of organisms that don’t sexually reproduce, or different species that hybridise with each other! Besides that, our knowledge and technology keep developing, which means that these concepts are still and probably always will be under debate (see for example Wheeler & Platnick, 2000).
While it’s easy to understand why spatially separated individuals of a species don’t get their genes flowing, there’s a bunch of less well-known mechanisms that prevent gene flow without geographical barriers. These are called Reproductive Isolating Barriers (RIBs), lead to speciation and can be divided in three main categories: pre-mating, pre-zygotic and post-zygotic barriers. Pre-mating RIBs mean that the male and female gamete never get to meet, for example because of a temporal mismatch or because the owners don’t recognise each other’s courting signals. For mating to turn into reproduction, we need a zygote to be formed. If there’s a pre-zygotic barrier such as a mismatch in physique or unsuccessful fertilisation, that means yet another barrier. If all this works and we have a hybrid zygote, it might simply not be able to survive in the environment that its parent organisms are well-adjusted to. Or it might have high mortality in no matter which environment, or is unable to produce offspring, all meaning we have a post-zygotic problem. Post-zygotic barriers are often caused by mismatches between the genes of the respective parents and are often found on the site of the heterogametic sex, i.e. the counterparts of humans that have XY instead of XX chromosomes. Different forms of reproductive isolation (RI) aren’t equally important, don’t necessarily occur equally fast in time and affect each other. These complications mean that experiments and technology in the lab and the field are incredibly important for our understanding of what’s (been) going on.
If we take a step back, we can at least say that evolutionary biologists have a pretty good understanding of what might be happening in individuals and their populations, and what processes and mechanisms can lead to the origination of a new species. There are several causes that can drive the processes that lead to speciation:
Ecological: two populations of one species adapt to different environments through natural selection;
Genetic conflict: a certain allele becomes very abundant but has negative effects on fertility, and another mutation, that repairs these effects, is incompatible with the “original” genetical set-up;
Sexual selection: sex A prefers certain visual, audio or other traits in sex B over other trait expressions, leading to certain varieties of certain traits always reproducing with each other instead of with other varieties in the species;
Reinforcement of RI: hybrids of two populations have lower survival or fertility meaning that they’ll be less successful reproducing than individuals from the two populations within themselves;
Polyploidy: genome-duplication can result in a new species within one generation when tetraploid individuals are unable to reproduce successfully with individuals from the original, diploid populations;
Hybrid speciation: the hybrid(s) of two “parent” species become genetically incompatible with them;
Genetic drift: a population that becomes fixed for a chromosomal rearrangement, a consequence of genetic drift, may no longer be compatible with the rest of the species.
The latter, speciation through genetic drift, occurs especially in small or highly-fluctuating populations. It is an important pillar under the founder effect: the start of a new population by a few individuals. If these few individuals have undergone changes due to genetic drift and get isolated from the rest of the population, they can come to form a new species if their genetic make-up is incompatible with the population of origin. This can be especially interesting at the edges of species ranges (Figure 2a), and there are descriptions of how this process can occur multiple times in a row, leading to a “wave” of changes through mutation (Figure 2b). If this leads to allele frequencies becoming relatively and unexpectedly high, this is called genetic “surfing” (Excoffier et al., 2009).
Figure 2. Allele frequencies a) during range expansion and b) during serial range expansion events. Derived from (Peischl et al., 2016).
While the founder effect is not fully understood or 100% supported by the literature, it’s clear that the geographical aspect that underlies it plays a role in many speciation events. If gene flow is completely blocked off by a geographical barrier, we speak of allopatric speciation. Note that the degree of geographic isolation depends on a geographic barrier, not on geographic distance, and that the degree to which it is an actual barrier also depends on the dispersal capacities of the species in question. Barriers can have their source in the environment, e.g. a new river divides two grasslands in two, or in a species itself, e.g. a few individuals manage to leave the mainland to start a population on an uninhabited island. Both can lead to allopatric speciation, but neither of these two processes has to be irreversible, as the river can fall dry again or the island species might expand their range to the mainland. In both situations, the populations of the mainland and the island can become sympatric. In such cases, we speak of secondary contact between the populations where, depending on the degree of reproductive isolation, gene flow is possible.
Sympatric speciation and the intermediate process, parapatric speciation, still have some gene flow going on. The former doesn’t depend on geographical barriers, but can depend on factors such as timing (e.g. flowering plants), local variation (e.g. soil characteristics) and behaviour (e.g. mating preferences). Sympatric speciation can reinforce itself if the allele in question is associated to another specific allele on another gene, which is rare but can occur when there’s a trait variety that works on both ecological divergence and reproductive isolation. Called a speciation trait, this trait can for example cause individuals with a specific feeding preference to only reproduce with each other. Other non-random mating patterns can lead to parapatric speciation. This leads to subpopulations continuing to interbreed with each other while getting more reproductively isolated from each other.
While the patterns of speciation are not easily understood and mapping them can become easier in collaboration with fields such as paleoarchaeology and geology, there is a lot of variation within the genome that’s left to be unravelled. The omics side of evolutionary biology can further the understanding of linkage equilibria, self-reinforcing processes and speciation genes and regions, to be able to understand what has happened in the past. Given the current pace of ecosystem degradation and destruction, we need this understanding of evolution to predict and, where possible, adapt to the future.
References
De Micco, V., & Aronne, G. (2012). Morpho-Anatomical Traits for Plant Adaptation to Drought. In R. Aroca (Ed.), Plant Responses to Drought Stress (pp. 37–61). Springer. https://doi.org/10.1007/978-3-642-32653-0_2
Excoffier, L., Foll, M., & Petit, R. J. (2009). Genetic consequences of range expansions. Annual Review of Ecology, Evolution, and Systematics, 40, 481–501. https://doi.org/10.1146/annurev.ecolsys.39.110707.173414
Futuyma, D., & Kirkpatrick, M. (2018). Evolution (4th ed.). Oxford University Press.
Laland, K., Matthews, B., & Feldman, M. W. (2016). An introduction to niche construction theory. Evolutionary Ecology, 30(2), 191–202. https://doi.org/10.1007/s10682-016-9821-z
Lenssen, J. P. M., Van Kleunen, M., Fischer, M., & De Kroon, H. (2004). Local adaptation of the clonal plant Ranunculus reptans to flooding along a small-scale gradient. Journal of Ecology, 92(4), 696–706. https://doi.org/10.1111/j.0022-0477.2004.00895.x
Love, S. J., Schweitzer, J. A., & Bailey, J. K. (2023). Climate‑driven convergent evolution in riparian ecosystems on sky islands. Scientific Reports, 1–9. https://doi.org/10.1038/s41598-023-29564-2
Peischl, S., Dupanloup, I., Bosshard, L., & Excoffier, L. (2016). Genetic surfing in human populations: from genes to genomes. Current Opinion in Genetics and Development, 41, 53–61. https://doi.org/10.1016/j.gde.2016.08.003
Simha, A., Pardo-De la Hoz, C. J., & Carley, L. N. (2022). Moving beyond the “Diversity Paradox”: The Limitations of Competition-Based Frameworks in Understanding Species Diversity. American Naturalist, 200(1), 89–100. https://doi.org/10.1086/720002
Wheeler, Q. D., & Platnick, N. I. (2000). The Phylogenetic Species Concept (sensu Wheeler and Platnick). In Q. D. Wheeler & R. Meier (Eds.), Species concepts and phylogenetic theory – a debate (pp. 55–68). Columbia University Press.
Two current members of NRRV, Larry Greenberg and Eva Bergman, and two former members, Johnny Norrgård and Pär Gustafsson, have recently published an overview of 15 years of research on the endemic, large-bodied population of landlocked River Klarälven-Lake Vänern population of Atlantic salmon (Salmo salar).
Large bodied adult Atlantic salmon from Lake Vänern
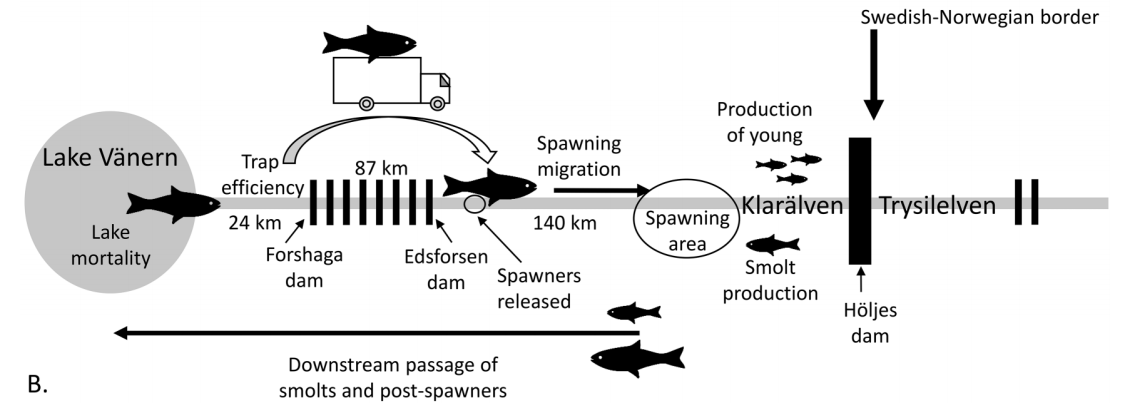
They highlight the major findings from studies of each of the salmon’s life stages and conclude that the Klarälven salmon population is below carrying capacity. Greenberg et al. (2021) suggest measures to increase the number of spawners and downstream passage success, and they also recommend habitat restoration to compensate for losses from, for example, former log-driving activities. They also discuss the ecological and legislative problems that need to be addressed if one wishes to re-establish salmon in Klarälven’s upper reaches in Norway. Managing, conserving and conducting research on this migratory salmonid population has been challenging not only because of the ecosystems’ large size, but also because there is more than one anthropomorphic stressor involved.
(B) Schematic diagram showing the different phases of the salmon life cycle in relation to the hydropower dams and the different studies undertaken in the River Klarälven – Lake Vänern ecosystem
A threespine stickleback (Gasterosteus aculeatus), a fascinating animal likely to be mentioned in both talks.
On Tuesday October 9, two seminars will be held at the biology department at Karlstad University.
Adaptive potential and evolutionary responses to climate change: Arctic char and threespine stickleback in Greenland – Michael Hansen, Professor, Aarhus University
Ecological genetics – What’s it about and how can we use it? – Karl Filipsson, NRRV PhD-student, Karlstad University
The seminars will be held in room 5F416 and start at 13:15. Everyone who wants to are welcome to attend the seminars.
Typically three spined sticklebacks have several spines in their dorsal and pelvic fins. In the ocean, the spines are important for predator defense in an environment with many large predators and few places to hide. For some landlocked populations the predation pressure can be lower or different (large predator fish may be lacking) and the spines may carry a cost instead of offering protection. In fact, several freshwater populations of sticklebacks in Europe and North America lack the pelvic spines – the result of evolution in a different environment. In the short film Making of the Fittest: Evolution of the Stickleback Fish, HHMI BioInteractive tells the story of the stickleback evolution and interviews researchers that track the genetic mechanism behind the change. Watch the film here.
On Tuesday, May 3, Anssi Laurila, from Uppsala University, will give a seminar titled “How to cope with environmental variation: a frog’s eye view on adaptive solutions”.
Here, Anssi Laurila describes his own research: “I work at the interface between ecology and evolutionary biology on factors allowing and maintaining phenotypic and genetic variation. Much of my research focuses on the roles of phenotypic plasticity and local adaptation in explaining phenotypic and genetic variation along environmental gradients. Within this theme, I am focusing on both large-scale (latitudinal) and small-scale climatic variation as well as other forms of environmental stress (acidification, disease) in explaining these patterns using amphibians and snails as models.”
The seminar will be given at 13:30 in room 5F416 on Karlstad University. Everyone is welcome to attend!
On Tuesday, April 18, Staffan Andersson from University of Gothenburg will give a seminar titled “Selection and constraints on plumage ornamentation in widowbirds and bishops (Euplectes spp)”.
The seminar will be given at 13:30 in room 5F416 on Karlstad University. Everyone is welcome to attend!